
Antibody type case, do you know why?
专栏:Industry news
发布日期:2018-01-11
阅读量:1805
Human beings live in a complex and ever-changing environment, and they are exposed to a variety of microorganisms every moment, and are invaded by some similar antigenic substances, causing the body to become ill. In order to resist these external intrusions and enable them to continue to survive, the human body must form hundreds of thousands, millions or even more specific antibodies to resist external antigens in order to protect themselves. We will elaborate and analyze the causes of antibody production and diversity.
1. Presentation of antigen
Antigen presenting cell (APC) antigen presentation is a complex process involving antigen uptake, processing and presentation [1]. The most important antigen presenting molecule is the major histocompatibility complex (MHC) [2]. MHC is divided into two broad categories: MHC-I and MHC-II, which present different sources of antigenic proteins, different ways of degrading antigens, and different lengths of peptides, which can present antigens to cytotoxic T cells (CD8 T, respectively). Cell) and helper T cells (CD4 T cell) [3,4].
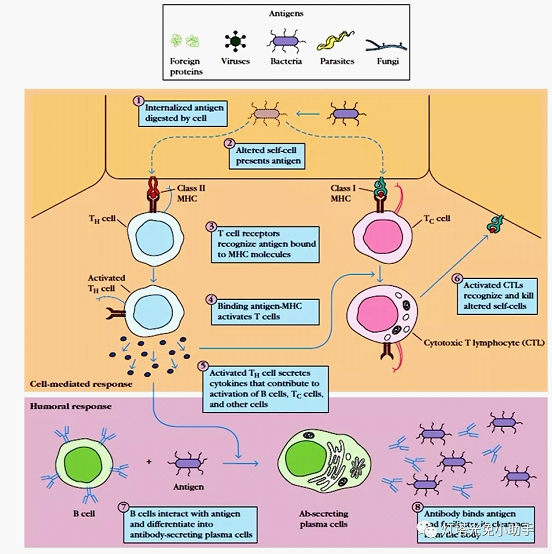
The antigen-presenting process is briefly described by presenting MHC-II as an exogenous protein.
The exogenous antigen is engulfed or swallowed by APC and is taken up into the phagosome, which fuses with the lysosome to form a phagolysosome. The antigen is degraded into small molecule polypeptides by proteolytic enzymes in the acidic environment of phagolysosomes, and the immunogenicity is called antigen peptide [3, 5]. The MHC class II molecules synthesized in the endoplasmic reticulum enter the Golgi apparatus and are carried by secretory vesicles. By fusion with phagolysosomes, the antigen peptides are combined with MHC class II molecules in the vesicles to form antigen peptide-MHC class II molecules. Complex. The resulting complex may be transported to the surface of the cell membrane with the participation of the Golgi complex, recognized by the T cell receptor (TCR) and presented to the TH cell [6]. A prerequisite for TCR recognition is the direct contact and interaction of the two cells. This intercellular interaction involves multiple molecules on the surface of APC and TH. In addition to TCR specifically recognizing a complex of polypeptide-MHC-II molecules simultaneously, certain adhesion molecules are also involved in the antigen presentation process. Activated TH cells secrete various cytokines for activation of B cells, Tc cells, etc., and produce specific antibodies that can scavenge antigens.
2. B cell differentiation and antibody production
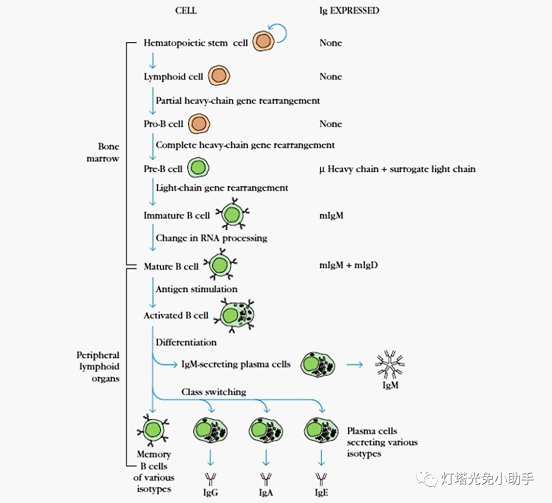
The differentiation process of mammalian B cells can be divided into five stages: pre-B cells, immature B cells, mature B cells, activated B cells and plasma cells. The differentiation of pre-B cells and immature B cells is antigen-independent, the differentiation process proceeds in the bone marrow, and the immature B cells begin to express membrane-bound IgM (mIgM). The antigen-dependent phase means that mature B cells (mIgM and mIgD are simultaneously expressed on the surface of the membrane) become activated B cells with the help of APC and TH cells after antigen stimulation (mIg levels are gradually decreased, secretory Ig is gradually increased, and Ig can occur) The class of gene heavy chains is transformed, and continues to differentiate into plasma cells that synthesize and secrete antibodies, and play a role in humoral immunity. This stage of differentiation is mainly carried out in peripheral immune organs.
Therefore, the antibody is present in a body fluid such as blood of a vertebrate, and the cell membrane surface of the B cell, and its main function is to effectively remove foreign matter such as microorganisms and parasites invading the body after binding to the antigen.
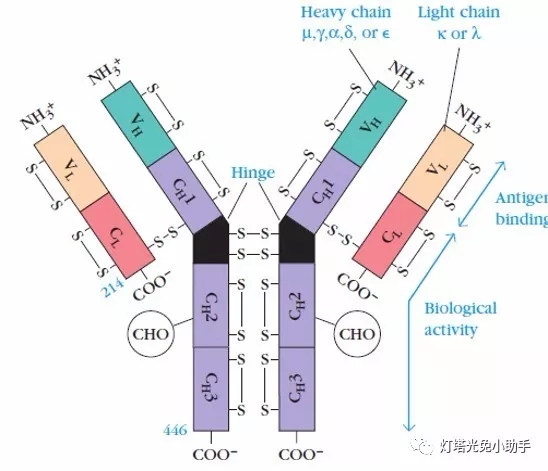
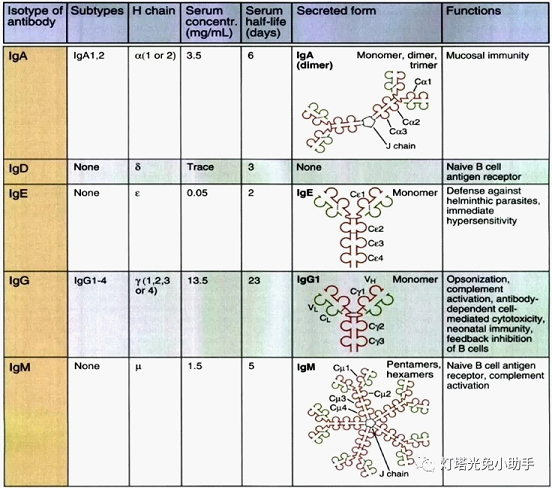
Antibodies are four symmetrical structure of a polypeptide chain, including 2 long, relative molecular weight of the larger the same heavy chain chain (H); Article 2 short, relative molecular weight smaller light chain of the same chain (L). By disulfide bond between chain and non covalent bond to form a monomer molecule made up of four polypeptide chain. Light chain a kappa and lambda predominate two, heavy chain with gamma, mu, alpha, epsilon and delta five [7], on the basis of different Ig heavy chain classification into IgM and IgG, IgA, IgD and IgE five classes.
The entire antibody molecule can be divided into two parts, a constant region and a variable region. In a given species, the constant regions of different antibody molecules have the same or nearly identical amino acid sequences. The variable region is located at the end of the two arms of the "Y", also known as the antigen-binding fragment (Fab). The two antigen binding sites on one antibody molecule are identical; the handle of the "Y" is called the crystalline fragment ( Crystalline fragment, FC).
3. Immunoglobulin diversity
The composition of antibodies is extremely complex and consists of thousands of diverse Ig molecules. These Ig molecules are similar and different in shape, size, structure, and composition and arrangement of amino acids.
3.1 Variable region heterogeneity
The genes in the constant region can determine the class and subclass of Ig molecules, which is one of the causes of Ig molecular diversity, but the main cause of the diversity of immunoglobulin molecules is the heterogeneity of the variable regions.
A small number of amino acid residues vary particularly strongly in the variable region, and the residue composition and arrangement order of these amino acids are more susceptible to mutation, which is called hypervariable region. The hypervariable region is located on the surface of the molecule and consists of up to 17 amino acid residues, ranging from 2 to 3. The hypervariable region is an antigen binding site complementary to an antigenic epitope, also known as complement-determining regions (CDRs) [8], whose amino acid sequence determines the specificity of binding of the antibody to the antigen antigen [9, 10]. The heavy and light chains each contain three CDRs that are circular in a three-dimensional structure and are specifically bound to the antigen on the surface of the antibody molecule.
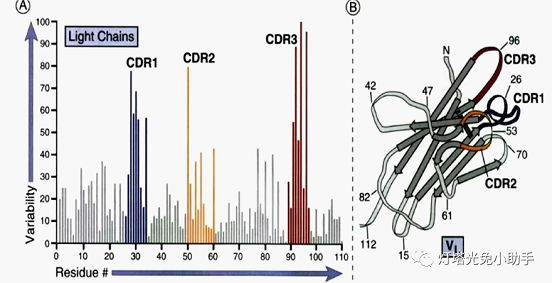
3.2 V (D) J recombination
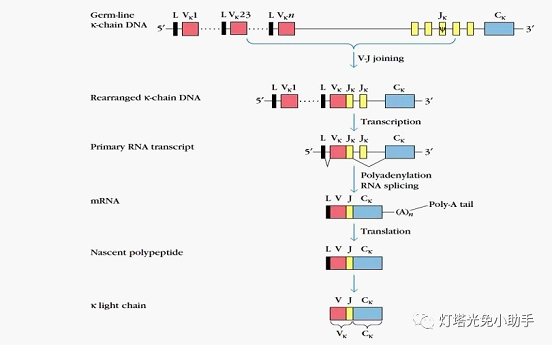
Antibodies by C, V, J L chain three gene cluster coding
H chain composed of C, V, D, J four genes encoding
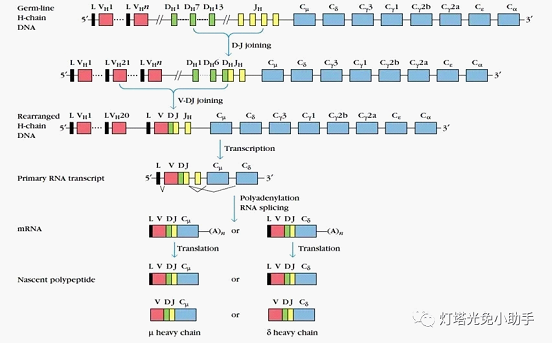
The human control immunoglobulin H chain is located on the long arm of chromosome 14, and the V gene fragment (VH), D gene fragment (DH), J gene fragment (JH), and C gene fragment encoding the variable region. (CH) Four types of gene fragments, of which 45 are VH, 23 are DH, 6 are JH, and 9 are CH [18]. The constant region on the H chain is controlled by the C gene fragment and is generally constant; the variable region on the H chain is controlled by three gene segments of VH, DH and JH. During the process of plasma cell maturation, random recombination can occur between the three gene segments of VH, JH and DH of heavy chain genes, and different combinations result in the production of heavy chain genes with different structures.
According to the somatic gene rearrangement theory, B cells begin rearrangement of the light chain (Lλ chain and Lκ chain) variable region genes after completion of rearrangement of the heavy chain variable region gene. The human control Lλ chain gene locus on the long arm of chromosome 22 has Vλ, Jλ and Cλ 3 gene fragments, of which 30 species are Vλ, 4 species are Jλ, and 4 species are Cλ. The constant region on the Lλ chain is controlled by the Cλ gene fragment and is generally constant; the variable region on the Lλ chain is controlled by the Vλ and Jλ2 gene segments. During the process of plasma cell maturation, random recombination can occur between the Vλ and Jλ gene fragments of the Lλ chain gene, and different combinations produce Lλ chain genes of different structures. Human locus control Lκ chain gene locus on the short arm of chromosome 2, there are Vκ, Jκ, Cκ 3 gene fragments, of which there are 40 species of Vκ, 5 species of Jκ, and only 1 species of Cκ [18].
These exons are under the action of recombinase (RE), through a variety of rearrangements, the peptide chain synthesized, and further L and H chain combinations, so that the final antibody type is very much. [12]. Antibody gene rearrangement occurs during lymphocyte differentiation [13].
3.3 category conversion
A B lymphocyte clone At the beginning of the differentiation process, immature B cells express only mIgM and mIgD, and both Ig have the same antigen binding region. After antigen-activated B cells, the Ig class expressed and secreted on the membrane is converted from IgM to other classes or subclasses of Ig [12] such as IgG, IgA, IgE. At this time, the VH gene fragment remains unchanged, and the rearrangement of the CH gene segment occurs, that is, the specificity of the binding antigen is the same, but the heavy chain type (constant region) changes. Thus, progeny cells of the same B cell can produce specific, different classes of antibodies against the same antigen. Local microenvironment and cytokines can influence and regulate the type conversion of immunoglobulins [14].
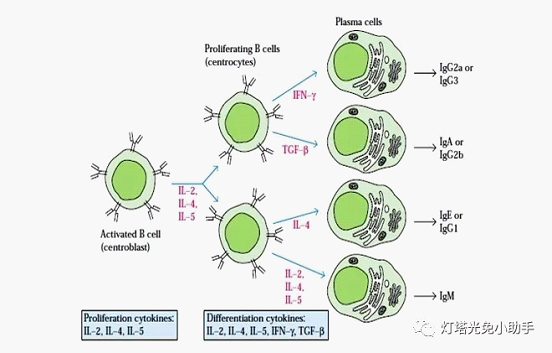
The mechanism by which cytokines regulate B cell Ig class switching may be: (1) stimulating the selective proliferation of clones of certain cells, and increasing the number of clonal cells secreting a specific class or subclass of antibodies, such as IL-5 and IL-6. In addition to regulation by isotype switching, IgA production can also selectively promote the differentiation of IgA-directed cells into IgA-secreting cells. (2) Inducing B cells to switch from secretory IgM to an isoform Ig by inducing recombination of two transition regions at specific positions.
3.4 Somatic cell high frequency mutation and affinity maturation of antibodies
After the B cells complete the rearrangement of the heavy chain gene and the light chain gene, the mature B cells undergo high frequency mutation after receiving antigen stimulation at the peripheral lymphoid germinal center, mainly in the base point mutation of the heavy chain region gene. Called somatic hypermutation [15]. High frequency mutations in somatic cells greatly increase the diversity of immunoglobulin genes, while also allowing the formation of antibody affinity to further mature.
The BCR affinities of various B cell clones produced after mutation are different, and B cells expressing high affinity antibodies on their surface will obtain strong survival signals in response to other cells, while B cells expressing low affinity antibodies gradually. Apoptosis [16]. The overall result is that the average affinity of the progeny B cells and the antibodies produced by them to the antigen is enhanced, called the affinity maturation of the antibody, so that the secreted antibody can protect the body from the foreign antigen more effectively. Invasion [17]. Only those B cells that express high-affinity antigen receptors can effectively bind antigen and proliferate in antigen-specific Th cells, producing high-affinity antibodies [17].
Under the guidance of the above diverse antibody genes, a variety of heavy chain protein molecules and light chain protein molecules have been formed. In addition, receptor editing, immunoglobulin class switching and other factors can greatly increase the diversity of immunoglobulin structures [18]. It can be said that there are many kinds of antigenic substances in nature, and how many kinds of antibodies can be formed by how many kinds of immunoglobulin molecules can be produced by the human body.
references
1. Mann, Elizabeth R. "Intestinalantigen-presenting cells in mucosal immune homeostasis: Crosstalk betweendendritic cells, macrophages and B-cells". World Journal ofGastroenterology. 20 (29): 9653.
2. Kambayashi, Taku;Laufer, Terri M. "Atypical MHC class II-expressing antigen-presentingcells: can anything replace a dendritic cell?". Nature Reviews Immunology.14 (11): 719–730.
3. Den Haan, Joke M.M.;Arens, Ramon; Zelm, Menno C. van. (2014). "The activation of the adaptiveimmune system: Cross-talk between antigen-presenting cells, T cells and Bcells". Immunology Letters. 162 (2): 103–112.
4. Hivroz, Claire;Chemin, Karine; Tourret, Marie; Bohineust, Armelle. "Crosstalk between TLymphocytes and Dendritic Cells". Critical Reviews™ in Immunology. 32 (2): 139–155.
5. Dalod, M.; Chelbi, R.;Malissen, B.; Lawrence, T. (2014). "Dendritic cell maturation: functionalspecialization through signaling specificity and transcriptionalprogramming". The EMBO Journal. 33 (10): 1104–1116.
6. Sadegh-Nasseri,Scheherazade; Chou, Chih-Ling; Hartman, Isamu Z.; Kim, AeRyon; Narayan, Kedar(2012). "How HLA-DM works: recognition of MHC II conformationalheterogeneity". Frontiers in Bioscience. 4: 1325–1332.
7. Charles Janeway(2001). Immunobiology (5th ed.). Garland Publishing.
8. Al-Lazikani B, LeskAM, Chothia C (1997). "Standard conformations for the canonical structuresof immunoglobulins". J Mol Biol. 273 (4): 927–948.
9. Woof J, Burton D(2004). "Human antibody-Fc receptor interactions illuminated by crystalstructures". Nat Rev Immunol. 4 (2): 89–99.
10. Heyman B (1996)."Complement and Fc-receptors in regulation of the antibody response".Immunol Lett. 54 (2–3): 195–199.
11. Nemazee D (2006)."Receptor editing in lymphocyte development and central tolerance".Nat Rev Immunol. 6 (10): 728–740.
12. Market E, PapavasiliouFN (2003). "V(D)J recombination and the evolution of the adaptive immunesystem". PLoS Biol. 1 (1): E16.
13. Market, Eleonora;Papavasiliou, F. Nina (2003). "V(D)J Recombination and the Evolution ofthe Adaptive Immune System". PLoS Biology. 1 (1): E16.
14. Stavnezer J, AmemiyaCT (2004). "Evolution of isotype switching". Semin. Immunol. 16 (4):257–275.
15. Diaz M, Casali P(2002). "Somatic immunoglobulin hypermutation". Curr Opin Immunol. 14(2): 235–240.
16 Or-Guil M, WittenbrinkN, Weiser AA, Schuchhardt J (2007). "Recirculation of germinal center Bcells: a multilevel selection strategy for antibody maturation". Immunol.Rev. 216: 130–41.
17. Neuberger M,Ehrenstein M, Rada C, Sale J, Batista F, Williams G, Milstein C (2000)."Memory in the B-cell compartment: antibody affinity maturation".Philos Trans R Soc Lond B Biol Sci. 355 (1395): 357–360.
18. 曹雪涛,熊思东,姚智.医学免疫学.北京:人民医学出版社,2013:77.